Ściana komórkowa
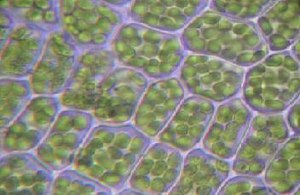
Ściana komórkowa komórki roślinnej
Ściana komórkowa – struktura otaczająca cytoplast komórek roślin, grzybów, bakterii, archeonów oraz części protistów. Zapewnia komórkom ochronę przed czynnikami fizycznymi, chemicznymi i biologicznymi, oraz decyduje o ich kształcie. U organizmów jednokomórkowych pełni funkcję szkieletu zewnętrznego. Zbudowana jest z wielu substancji, a skład jest zależny od przynależności taksonomicznej. Wspólną cechą jest występowanie w ścianie komórkowej biopolimerów (celulozy, chityny, mureiny), tworzących jej szkielet. Poza składnikami szkieletowymi w skład ściany komórkowej mogą wchodzić inne polisacharydy, lipoproteiny, lipopolisacharydy lub inne lipidy. Składniki zapewniają strukturze zarówno znaczną sztywność, jak i duży stopień elastyczności[1].
Leżąca na zewnątrz błony komórkowej ściana komórkowa to pierwsza struktura komórkowa poznana dzięki obserwacjom mikroskopowym. Jej istnienie zostało stwierdzone przez Roberta Hooke’a w roku 1663[2].
Polisacharydy ściany komórkowej roślin są składnikiem tkanin, papieru, drewna budowlanego, folii, środków zagęszczających i innych, a celuloza jest najbardziej rozpowszechnionym i użytecznym biopolimerem na Ziemi[3]. Przewiduje się, że wykorzystanie substancji ścian komórkowych będących głównym składnikiem słomy zbóż i innych pozostałości roślinnych będzie rosło w wyniku stosowania odnawialnych źródeł energii oraz wytwarzania materiałów kompozytowych[4].
Spis treści
1 Funkcje
2 Budowa i skład
2.1 Bakterie
2.2 Archeany
2.3 Grzyby
2.4 Protisty
2.5 Rośliny
2.5.1 Typy ścian w komórkach roślinnych
3 Biogeneza
4 Ewolucja
5 Przypisy
Funkcje |
Ściana komórkowa nadaje kształt i sztywność komórce. Jest przepuszczalna dla soli mineralnych i innych substancji drobnocząsteczkowych dlatego utrzymywanie kształtu jest powiązane z ciśnieniem wewnętrznym, czyli turgorem związanym z selektywną przepuszczalnością błony komórkowej[1]. Struktura pełni również funkcje ochronne, a u organizmów pasożytniczych dzięki obecności enzymów hydrolitycznych i cząstek toksycznych umożliwia inwazję[5].
U roślin będących organizmami tkankowymi ściany komórkowe są częścią apoplastu, przedziału zewnątrzkomórkowego przez który transportowana jest znaczna ilość wody i substancji w niej rozpuszczonych. Struktura ścian komórkowych pozwala regulować transport zachodzący w apoplaście. Za największe cząsteczki białek, które mogą przemieszczać się w obrębie porów ścian uznaje się cząstki o masie 10–50 kDa. Komórki niektórych tkanek poprzez depozycję substancji hydrofobowych stają się nieprzepuszczalne dla wody. Tak jest na przykład w komórkach endodermy. Konieczność przejścia wody i związków w niej rozpuszczonych przez błonę komórkową pozwala ściśle kontrolować transport tych substancji. Połączenie cytoszkieletu ze ścianą komórkową ułatwia rozmieszczenie organelli wewnątrz komórki oraz ich ruchy. Możliwe jest także przeniesienie sygnałów chemicznych oraz mechanicznych z powierzchni komórki do wnętrza jądra. Wiele znanych cząsteczek sygnałowych, na przykład tlenek azotu i reaktywne formy tlenu, generowanych jest w ścianie komórkowej. Umożliwiają one rekcje komórek na stres powodowany przez czynniki biotyczne i abiotyczne. Fragmenty oligosacharydowe oraz glikoproteinowe mogą pełnić funkcję elicytorów. Bodźce ze środowiska wpływają na strukturę ścian komórkowych tkanek powodując między innymi wykształcanie drewna reakcyjnego, kompresyjnego i tensyjnego. W tkankach spichrzowych hemicelulozy są ważnym materiałem zapasowym. Podczas kiełkowania zgromadzone w bielmie oraz liścieniach hemicelulozy są hydrolizowane, a powstające cukry służą jako źródło energii[2].
Budowa i skład |
Bakterie |

Ściana komórkowa bakterii tworzy woreczek
Ściana komórkowa bakterii tworzy po zewnętrznej stronie błony komórkowej woreczek (sacculus). Z zewnątrz pokryta jest otoczką (błoną zewnętrzną). Woreczek ma grubość 2–10 nm u bakterii Gram-ujemnych i 15–50 nm u bakterii Gram-dodatnich[6]. U tych pierwszych ściana komórkowa składa się z tylko jednej warstwy mureiny, zaś u drugich z około 40 warstw[1]. Podstawowym składnikiem bakteryjnej ściany komórkowej jest polimer kwasu N-acetylomuraminowego i N-acetyloglukozoaminy. Związki te – połączone wiązaniem β-1,4 – tworzą łańcuchy o długości kilkudziesięciu monomerów. Biopolimer nazywany mureiną lub peptydoglikanem jest dodatkowo usieciowany dzięki połączeniu łańcuchów wielocukrów powstałym przez przyłączenie krótkich polipeptydów do reszt kwasy muraminowego. Połączenia pomiędzy peptydami powstają powstają przez kwas diaminopimelinowy, lizynę, lub mostka, składającego się z kilku reszt aminokwasowych (u gronkowca złocistego mostek tworzy pięć reszt glicyny)[6]. Mureina stanowi u bakterii Gram-dodatnich od 30% do 70% suchej masy. W składzie ściany tych bakterii stwierdza się również obecność kwasów tejchojowych oraz pewne ilości białek. U bakterii Gram-ujemnych mureina stanowi mniej niż 10% suchej masy ściany komórkowej. Łańcuchy polisacharydowe nie są połączone mostkami międzypeptydowymi, ani z udziałem lizyny. Połączenia powstają jedynie przez kwas diaminopimelinowy. Poza tworzącą szkielet ściany komórkowej mureiną w jej skład wchodzą znaczne ilości lipoprotein, lipopolisacharydów oraz innych lipidów. Część tych związków jest kowalencyjnie związana z mureiną[1]
Archeany |
U archeanów ściana komórkowa jest cieńsza niż u bakterii. Jej skład jest zróżnicowany w zależności od przynależności taksonomicznej. Methanobacterium mają ścianę komórkową, w której podstawowym składnikiem jest pseudomureina. Kwas N-acetylomuraminowy jest w niej zastąpiony przez kwas N-acetylotalozoaminouronowy. U Methanosarcina elementy szkieletowe zbudowane są z metanochondroityny, biopolimeru powstającego z N-acetyloglukozaaminy lub galaktozoaminy oraz kwasów uronowych i krótkich peptydów. W skład peptydów wchodzą reszty lizyny, alaniny i glutaminy. Ściany komórkowe archeanów mogą być zbudowane także z innych heteropolisacharydów[6].
U niektórych grup struktura tworząca osłonkę nie przypomina typowej ściany komórkowej. U Thermoplasma łańcuchy cukrowe są elementem lipoglikanów i lipoprotein wbudowanych w błonę komórkową. Wystając poza nią tworzą barierę między komórką a środowiskiem. Łańcuchy te są bogate w mannozę. Methanospirillum i Methanococcus wytwarzają jedynie luźną i cienką osłonę białkową. Osłona chroniąca przez wpływem środowiska określana jest jako warstwa S[6].
Grzyby |
Struktura i powstawanie ściany komórkowej u grzybów jest charakterystyczne dla tej grupy organizmów[7]. Elementy szkieletowe mogą być zbudowane z chityny, chitozanu, kompleksów chityna-glukany. W skład macierzy wchodzą glukany oraz glikoproteiny[8]. Ściana komórkowa ma budowę warstwową. W warstwie najbardziej wewnętrznej występują mikrofibryle chitynowe i białka. W warstwach pośrednich wchodzą polisacharydy, białka i glikoproteiny. Warstwa najbardziej zewnętrzna składa zbudowana jest z bezpostaciowych glukanów[9].
Protisty |

Pancerzyki okrzemek są szczególnym rodzajem ściany komórkowej
U Hyphochytridiomycetes składnikami szkieletowymi ściany komórkowej są zarówno chityna, jak i celuloza[8]. Cechą charakterystyczną Oomycetes jest ściana komórkowa zawierająca celulozę, chociaż u nielicznych (Leptomitales) w skład ściany komórkowej wchodzi także chityna[10]. Badania na Saprolegnia monoica wskazują, że chityna u tego gatunku ma postać globularną i nie tworzy struktur o charakterze mikrofibryli[11]. U innych protistów zaliczanych do glonów w skład ściany komórkowej wchodzi celuloza. Szczególnym przypadkiem są okrzemki, które wytwarzają pancerzyki, których głównym składnikiem jest krzemionka[12].
Rośliny |

Ściana komórkowa komórki roślinnej zaznaczona na zielono

Budowa pierwotnej ściany komórkowej u roślin
Grubość ściany komórkowych roślin zwykle mieści się w granicach od około 0,1 do 10 μm[13].
U roślin elementy szkieletowe ściany komórkowej zbudowane są z celulozy. Matriks (podłoże) składa się z hemiceluloz i pektyn. Hemicelulozy to głównie polimery zbudowane z reszt kwasu glukuronowego oraz pentoz takich jak ksyloza i arabinoza lub w drugim wariancie pozbawione reszt kwasu glukuronowego, składające się jedynie z pentoz (ksylozy i arabinozy) oraz heksoz (mannoza, glukoza, galaktoza). Zależnie od dominującego składnika cukrowego takie hemicelulozy nazywane są ksylanami, mannanami, galaktanami. W skład matriks wchodzi także niewielka ilość białek. Są wśród nich metyloesterazy pektyn oraz hemicelulazy, uczestniczące w syntezie składników ściany, ekspansyna wpływająca na rozciągliwość struktury, ekstensyna, będąca białkiem strukturalnym. Celuloza tworząca elementy szkieletowe może mieć postać krystaliczną albo amorficzną. Z celulozy krystalicznej tworzą się regiony micelarne. Są one przedzielone rejonami międzymicelarnymi o średnicy około 1 nm. Pojedyncza cząsteczka celulozy składa się z 1000–25 000 reszt glukozowych. Długość łańcucha jest zależna od gatunku rośliny, jest także różna dla ściany komórkowej pierwotnej i wtórnej. Cząsteczki celulozy tworzą struktury wyższego rzędu nazywane mikrofibrylami i makrofibrylami. Pojedyncza fibryla elementarna składa się zwykle z 36 ułożonych równolegle cząsteczek celulozy. Mikrofibryle tworzone są przez 3–4 fibryle elementarne, zaś makrofibryle składają się z 200–400 mikrofibryl. Struktury te są widoczne pod mikroskopem świetlnym i mają średnicę około 0,2–0,5 μm[14].
W komórkach roślinnych mogą występować dwie warstwy ściany komórkowej. Młode komórki wykształcają jedynie cienką ścianę pierwotną. W jej skład wchodzą wyłącznie elementy szkieletowe z celulozy oraz macierz z hemiceluloz, pektyn oraz białek. Zawartość celulozy wynosi około 20%. Taka ściana komórkowa jest cienka i bardzo elastyczna. Komórki, które przestały się powiększać, wytwarzają kolejną warstwę ściany komórkowej nazywaną ścianą wtórną. Nowe warstwy nakładane są od strony protoplastu. Zwykle w trakcie pogrubiania ściany komórkowej nakładane są trzy warstwy składające się na ścianę wtórną. Ich podstawowe składniki nie różnią się od substancji budujących ścianę pierwotną, jednak zawartość celulozy może być znacznie większa niż w ścianie pierwotnej. Zwykle wynosi około 60%, a we włoskach nasion bawełny może przekraczać nawet 90%[9].
Wtórna ściana komórkowa może ulegać modyfikacjom, które polegają na wnikaniu substancji między elementy szkieletowe (inkrustacja) lub odkładaniu dodatkowych związków chemicznych na powierzchni ściany (adkrustacja). Jednym z głównych związków prowadzanych do ściany komórek roślinnych w wyniku inkrustacji jest lignina. Efektem lignifikacji jest nieprzepuszczalność ściany dla wody oraz zwiększenie jej odporności chemicznej oraz mechanicznej[14]. U drzew w skład wtórnej ściany komórkowej wchodzi znaczna ilość ksylanów[2]. W komórkach tkanki korkowej pomiędzy pierwotną a wtórną ścianą komórkową odkładana jest suberyna. Proces określany jest jako korkowacenie[9]. Proces ten zachodzi głównie w korku, ale także np. w komórkach egzodermy i endodermy.
Inkrustacja może też prowadzić do wzbogacenia ściany komórkowej w substancje mineralne najczęściej krzemionką SiO2 (np. turzyce, skrzypy) lub węglanem wapnia CaCO3 (ramienice). W efekcie mineralizacji ściany stają się twarde, sztywne, a zarazem łamliwe[14].
Wtórne ściany komórkowe mogą osiągać znaczną grubość. W przypadku komórek sklerenchymy wypełniają niemal całe wnętrze komórki. Znaczną grubość osiągają również ściany cewek i naczyń. Szczególny rodzaj ściany komórkowej otacza ziarna pyłku. Warstwa zewnętrzna odpowiadająca pierwotnej ścianie komórkowej określana jest jako intyna, warstwa odpowiadająca ścianie wtórnej to egzyna. Odkładane są w niej znaczne ilości sporopoleniny, substancji o wyjątkowej odporności chemicznej. Wtórnym przekształceniom zachodzącym w obrębie pierwotnej ściany komórkowej ulegają komórki epidermy[2]. W wyniku kutynizacji, odkłania kutyny pomiędzy elementami szkieletowymi zewnętrzna ściana komórkowa staje się trudno przepuszczalna dla wody, chroniąc roślinę przez jej utratą. Dodatkowo, w procesie kutykularyzacji, na powierzchni ściany mogą odkładać się woski i kutyna tworząc kutykulę[9].
Ściany komórkowe są jednym z głównym magazynów wapnia u roślin[13].
Typy ścian w komórkach roślinnych |
Ze względu na skład wyróżniane są ściany typu I i typu II. W skład obu typów ścian wchodzą wszystkie związki występujące w pierwotnej ścianie komórkowej, jednak proporcje między nimi mogą znacznie się różnić. W ścianach typu I, które są obecne u roślin dwuliściennych i części roślin jednoliściennych celuloza i hemicelulozy występują w porównywalnej ilości. Głównym glukanem jest fukogalaktoksyloglukan. W ścianie występują niewielkie ilości związków fenolowych. Wyjątkowo u Lamiales i Solanales dominuje hemiceluloza – ababinoksyloglukan. W ścianach typu II, występujących u traw i roślin z nimi spokrewnionymi (rośliny komelinopodobne), dominującymi polisacharydami są hemicelulozy, głównie glukuronoarabinoksylany. Zawartość pektyn i białek jest niewielka., a podstawowym składnikiem stabilizującym strukturę są związki fenylopropanoidowe[2].
Biogeneza |
W komórce roślin wyższych tworzenie ściany komórkowej rozpoczyna się w cytokinezie. W pierwszym etapie w płaszczyźnie równikowej dzielącej się komórki formują się mikrotubule i mikrofilamenty. Elementy te układają się równolegle do siebie. Dochodzi do zawiązania fragmoplastu. Mikrotubule i mikrofilamenty kontrolują kierunek transportu oraz proces łączenia się pęcherzyków z cząsteczkami kalozy, hemiceluloz i pektyn. Pęcherzyki te wytwarzane są przez aparaty Golgiego. Pęcherzyki zlewają się ze sobą, tworząc trójwarstwową przegrodę w obrębie dzielącej się komórki, zwanej przegrodą pierwotną. Warstwa środkowa otoczona jest 2 błonami plazmatycznymi. Ze zlewających się pęcherzyków powstaje początkowo struktura cystern połączonych rureczkami, a następnie podziurawiona przegroda pierwotna. Rozrasta się ona odśrodkowo aż do momentu zetknięcia z błoną komórkową. Następuje połączenie błon. Kaloza wchodząca w skład przegrody pierwotnej ulega hydrolizie a pektyny zespalają przeciwne końce ściany komórkowej tworząc blaszkę środkową. W kolejnym etapie następuje wytworzeni ściany pierwotnej, a po ustaniu wzrostu komórki powstaje ściana wtórna[14]. Niezbędne do powstania macierzy polisacharydy wytwarzane są w aparatach Golgiego i transportowane do ściany poprzez egzocytozę. Następnie następuje ich wbudowanie pomiędzy mikrofibryle celulozowe[15]. Celulozowe elementy szkieletowe są wytwarzane w błonie komórkowej i bezpośrednio osadzane w docelowej lokalizacji[16].
Pomimo znacznej sztywności ściany komórkowej możliwy jest wzrost komórki z już wykształconą struktura. W trakcie wzrostu dochodzi do rozluźnienie elementów. W procesie tym biorą udział ekstensyny, endotransglikozylaza ksyloglukanu oraz endo-1,4-beta-D-glukanaza[17].
Ewolucja |
Współczesna wiedza o strukturze ściany komórkowej i ewolucji jej struktury jest ograniczona. Bez wątpienia obecność ściany komórkowej miało duże znaczenie w procesie kolonizacji środowiska lądowego przez rośliny, które nastąpiło około 470 milionów lat temu. Badanie ewolucji ściany komórkowej opiera się przede wszystkim na analizie jej różnorodności u poszczególnych grup organizmów. Badania te pozwalają na powiązanie morfologii z rolą fizjologiczną. Geny i enzymy zaangażowane w syntezę poszczególnych składników ściany komórkowej nie są jeszcze w pełni znane[18].
Szeroko rozpowszechniona w królestwie roślin celuloza występuje także u glonów, bakterii w tym sinic oraz osłonic. Geny syntazy celulozy wydają się być wspólne dla Chlorophyta i zielenic, krasnorostów i brunatnic. Enzym syntetyzujący mannany i glukomannany (CslAs) jest prawdopodobnie nieobecny u zielenic. Doświadczenia wykazały jednak obecność mannanów w ścianach komórkowych niektórych krasnorostów. Ksylany występują powszechnie u roślin naczyniowych, glewików, zielenic i krasnorostów. Geny niezbędne do ich syntezy powstały więc zapewne jeszcze przed rozdzieleniem zielenic i krasnorostów. Jednym z najstarszych składników ściany komórkowej są prawdopodobnie białka AGP (od ang. arabinogalactan proteins), wykryte zarówno u roślin lądowych, jak u zielenic, krasnorostów i brunatnic[19]. Dotychczas zebrane dane wskazują, że od momentu pojawienia się roślin lądowych zmiany w składzie polisacharydów nie zachodziły, chociaż kilka polisacharydów – w tym ramnogalakturonan II (RG II) – wydaje się być innowacją wprowadzoną już podczas ewolucji roślin telomowych[20].
Przypisy |
↑ abcd Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydawnictwo Naukowe PWN, 2004, s. 71–87. ISBN 83-01-13999-4.
↑ abcde Przemysław Wojtaszek: Ściana komórkowa. W: Przemysław Wojtaszek, Adam Woźny, Lech Ratajczak: Biologia komórki roślinnej. Warszawa: Wydawnictwo Naukowe PWN, 2008, s. 227–269. ISBN 978-83-01-14869-0. OCLC 749661223.
↑ D.J. Cosgrove. Growth of the plant cell wall. „Nat Rev Mol Cell Biol”. 6 (11), s. 850–861, Nov 2005. DOI: 10.1038/nrm1746. PMID: 16261190.
↑ N. Farrokhi, R.A. Burton, L. Brownfield, M. Hrmova i inni. Plant cell wall biosynthesis: genetic, biochemical and functional genomics approaches to the identification of key genes. „Plant Biotechnol J”. 4 (2), s. 145–167, Mar 2006. DOI: 10.1111/j.1467-7652.2005.00169.x. PMID: 17177793.
↑ Jean-Paul Latgé. The cell wall: a carbohydrate armour for the fungal cell. „Molecular Microbiology”. 66 (2), s. 279–290, 2007. DOI: 10.1111/j.1365-2958.2007.05872.x. ISSN 0950-382X (ang.).
↑ abcd Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydawnictwo Naukowe PWN, 2005, s. 89–95. ISBN 83-01-14378-9.
↑ Shaun M. Bowman, Stephen J. Free. The structure and synthesis of the fungal cell wall. „BioEssays”. 28 (8), s. 799–808, 2006. DOI: 10.1002/bies.20441. ISSN 0265-9247 (ang.).
↑ ab G.W. Gooday. Cell Walls. „The Growing Fungus”, s. 43–62, 1995. DOI: 10.1007/978-0-585-27576-5_3 (ang.).
↑ abcd Szweykowska Alicja, Szweykowski Jerzy: Botanika t.1 Morfologia. Warszawa: Wydawnictwo Naukowe PWN, 2003, s. 59–70. ISBN 83-01-13953-6.
↑ Szweykowska Alicja, Szweykowski Jerzy: Botanika t.2 Systematyka. Warszawa: Wydawnictwo Naukowe PWN, 1993, s. 481. ISBN 83-01-10950-5.
↑ Vincent Girard. Characterization of chitin and chitin synthase from the cellulosic cell wall fungus Saprolegnia monoica. „Experimental Mycology”, 1991. DOI: 10.1016/0147-5975(92)90037-R (ang.).
↑ David S Domozych. Algal Cell Walls. „eLS”, 2011. DOI: 10.1002/9780470015902.a0000315.pub3 (ang.).
↑ ab Przemysław Wojtaszek, Adam Woźny, Lech Ratajczak: Biologia komórki roślinnej. Tom 1. Struktura. Warszawa: Wydawnictwo Naukowe PWN, 2006, s. 4-9. ISBN 978-83-01-14838-6.
↑ abcd Andrzej Tretyn: Podstawy strukturalno-funkcjonalne komórki roślinnej W: Fizjologia roślin (red. Kopcewicz Jan, Lewak Stanisław). Warszawa: Wydawnictwo Naukowe PWN, 2002, s. 22–88. ISBN 83-01-13753-3.
↑ A.P. Sandhu, G.S. Randhawa, K.S. Dhugga. Plant cell wall matrix polysaccharide biosynthesis. „Mol Plant”. 2 (5), s. 840–850, Sep 2009. DOI: 10.1093/mp/ssp056. PMID: 19825661.
↑ D.M. Gibeaut, N.C. Carpita. Biosynthesis of plant cell wall polysaccharides. „FASEB J”. 8 (12), s. 904–915, Sep 1994. PMID: 8088456.
↑ D.J. Cosgrove. Expansive growth of plant cell walls. „Plant Physiol Biochem”. 38 (1–2), s. 109–124, 2000. PMID: 11543185.
↑ J.U. Fangel, P. Ulvskov, J.P. Knox, M.D. Mikkelsen i inni. Cell wall evolution and diversity. „Front Plant Sci”. 3, s. 152, 2012. DOI: 10.3389/fpls.2012.00152. PMID: 22783271.
↑ Z.A. Popper, M.G. Tuohy. Beyond the green: understanding the evolutionary puzzle of plant and algal cell walls. „Plant Physiol”. 153 (2), s. 373–383, Jun 2010. DOI: 10.1104/pp.110.158055. PMID: 20421458.
↑ I. Sørensen, D. Domozych, W.G. Willats. How have plant cell walls evolved?. „Plant Physiol”. 153 (2), s. 366–372, Jun 2010. DOI: 10.1104/pp.110.154427. PMID: 20431088.


